|
Úvod do sportovní fyziologie |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
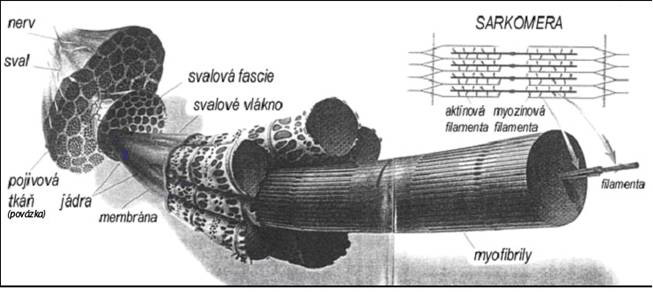
Stavba svalu
Lidské tělo obsahuje asi 660 svalů. Sval tvoří z cca 70% voda, asi 20% představují proteiny. Základní jednotkou kosterního svalu jsou svalová vlákna, podlouhlé buňky cylindrického tvaru navzájem spojené vazivem do svalových snopců (fascií), jejichž svazky tvoří sval, připevněný na kosti šlachou. Každé svalové vlákno je složeno z membrány (sarkolemy), vnitřní tekuté cytoplazmy (sarkoplazmy, cytosolu), buněčných organel (mitochondrie, ribozomy, sarkoplazmatické retikulum aj.) a tisíců myofibril, podélných vlákének, které vyplňují cytoplazmu uvnitř buňky. Největší svalová vlákna v lidském těle jsou až 30 cm dlouhá a 0,05-0,15 mm široká a obsahují tisíce jader.
Myofibrily jsou uvnitř položeny v podélných svazcích a jsou stejně dlouhé jako vlákno. Sestávají z podélných řetězců tzv. sarkomer, jež obsahují dva kontraktilní proteiny (filamenta), myozin a aktin, a ještě několik dalších proteinů, které jejich činnost regulují (např. troponin nebo elastický titin). V klidu jsou silnější myozinová filamenta obklopena slabšími aktinovými filamenty a navzájem jsou jen ve slabém kontaktu. Svalový stah (svalová kontrakce) se uskutečňuje pomocí tzv. příčných můstků (myozinových hlav) na myozinových filamentech. Dochází k tomu vlivem nervového impulsu z motorických nervů (motoneuronů), jež jsou situovány v míše nebo mozkovém kmeni. Jeden motorický nerv ovládá až stovky svalových vláken prostřednictvím tzv. motorických plotének na povrchu svalového vlákna. Nervový impuls vycházející z motorické ploténky nejprve vypudí z trubicovitých organel okolo buněčného jádra (tzv. sarkoplasmatického/endoplasmatického retikula) ionty vápníku (Ca+). Jejich působením se uvolňují troponinové "zámky" mezi filamenty, myozinové hlavy se navážou na aktin, sklopí se a vtahují se mezi filamenta aktinu. Svalová vlákna ovládaná jedním motorickým nervem dohromady tvoří samostatnou motorickou jednotku a jsou ve svalu navzájem promíchána mezi vlákny jiných motorických jednotek, což umožňuje lepší adaptaci svalu na svalovou práci.
Energii pro pohyb příčných můstků poskytuje adenosintrifosfát (ATP). Na každý pohyb příčného můstku je nutno rozštěpit hydrolýzou (reakcí s vodou) jednu molekulu ATP. Štěpením ATP vzniká adenosindifosfát (ADP) a organický fosfor (P). U dospělého člověka existují příčné můstky v několika hlavních variantách (izoformách), které závisí na typu motorického nervu, jimž je sval ovládán. Izoformy určují vlastnosti svalového vlákna a rychlost jeho smršťování. V zásadě jsou rozlišovány tři základní izoformy I, IIa a IIb (IIx).
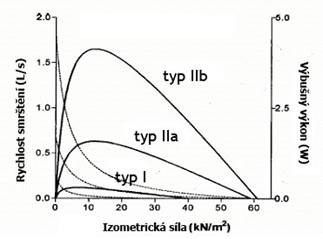
Podle nich se svalová vlákna dělí tři hlavní typy I, IIa a IIb: Pomalá (oxidativní) vlákna typu I jsou nezbytná pro vytrvalostní, aerobní svalovou práci (tj. dlouhodobou, méně intenzivní práci probíhající za přístupu kyslíku). Smršťují se sice pomalu (70-140 milisekund), avšak využívají energii ATP efektivněji a jsou typické velkou hustotou prokrvení. Mají malý průřez, dlouhé sarkomery, obsahují málo glykogenu (zásobní forma glukózy), málo enzymů účastných v glykolytických (anaerobních) reakcích, ale zato mají vysoký obsah oxidativních (aerobních) enzymů, které hrají roli v oxidativních reakcích (Krebsův cyklus), a vysoké zásoby triacylglycerolů (triglyceridů; zásobní formy tuků). Obsahují rovněž velké množství proteinu myoglobinu, v němž jsou uloženy pohotové zásoby kyslíku pro intenzívní svalovou práci.
Vlákna rychlá (typ II) se dělí na pomalejší typ IIa (oxidativně-glykolytický), jenž má i určitý aerobní potenciál, a rychlejší typ IIb či IIx (glykolytický), který je důležitý pro anaerobní sporty, kde dominuje explozívní energie, jako jsou např. krátké sprinty či skoky. Vlákna typu IIa představují jakýsi přechod mezi vlákny I a IIb; mají velký průřez, kratší sarkomery, střední obsah myoglobinu a mitochondrií, méně husté prokrvení, poměrně velké zásoby glykogenu i kreatinfosfátu, málo triacylglycerolů a střední rychlost smrštění (50-100 ms). Aktivita oxidativních i glykolytických enzymů je srovnatelně vysoká. Vlákna typu IIb mají největší dynamickou sílu ze všech tří typů, ale nízký obsah mitochondrií i myoglobinu a malé prokrvení. Vlivem menšího množství cytoplazmy je jejich průřez menší než u vláken typu IIa a obvykle jen nepatrně větší než u vláken typu I. Mají velké zásoby kreatinfosfátu a glykogenu, avšak málo triacylglycerolů. Smršťují se asi 4krát rychleji než vlákna typu I (20-50 ms). Aktivita glykolytických enzymů je vysoká, oxidativní enzymy jsou málo činné. Vlákna I a IIa jsou označována také jako vlákna červená. Toto zbarvení jim dodává myoglobin. Rychlá vlákna IIb se označují jako tzv. vlákna bílá. Vedle těchto tří základních typů svalových vláken jsou známy ještě čtyři další s hybridními fyziologickými charakteristikami (Ic, IIc, IIac, IIab), ale ty nejsou za běžných okolností rozlišovány.
Průřez a metabolismus svalových vláken lze do jisté míry ovlivnit sportovním tréninkem. Jejich složení – a tedy i rychlost kontrakce - je však možno změnit pouze částečně. Kupříkladu vlákna typu IIb se velmi plasticky přizpůsobují druhu tělesného zatížení a při dlouhodobém intenzivním tréninku s určitým podílem aerobní složky (vytrvalostním i běžném posilovacím) se prakticky kompletně konvertují na vlákna typu IIa. Je to zřejmě způsobeno tím, že extrémní anaerobní charakter vláken typu IIb se v běžném životě i při sportovním tréninku uplatňuje pouze v menší míře a nedostatek některých buněčných organel účastných při anabolických reakcích (mitochondrie) může být dokonce překážkou zlepšování výkonnosti. Řada výzkumů například jasně dokládá, že nárůst svalové síly a objemu výrazně závisí na přeměně IIb > IIa. Proto je výhodné, aby se vlákna IIb mohla podle potřeby přizpůsobovat pracovním požadavkům s podílem aerobní složky.
Ačkoli konverze typu II > I, I > II byly v některých studiích doloženy, je zřejmé, že probíhají v podstatně menším rozsahu nebo během velmi dlouhé doby a výsledky jsou často rozporuplné. Výrazné přeměny izoforem byly zatím doloženy pouze při dlouhodobém znehybnění (I > II) nebo při pokusech s elektrickou stimulací svalů laboratorních zvířat. Navíc dosud neexistuje dostatečné množství dlouhodobých studií, které by nám mohly říci, do jaké míry byl poměr rychlých a pomalých vláken dán elitním sportovcům „do kolébky“ nebo jak byl ovlivněn dlouholetým tréninkem. U vytrvalců byla nicméně objevena závislost mezi počtem let vytrvalostního tréninku a poměrem pomalých vláken.
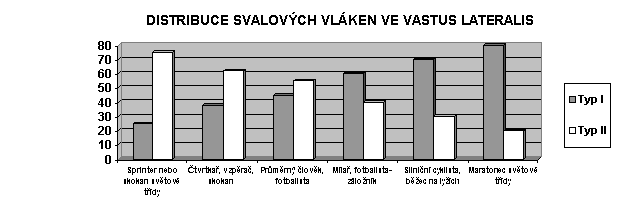
Poměr počtu rychlých a pomalých vláken je ve většině svalů přibližně rovnoměrný (50% : 50%), přičemž vlákna IIb tvoří ze všech tří typů nejmenší podíl (obvykle asi 10-20%). Vzhledem ke své větší velikosti však rychlá vlákna II zabírají více než polovinu celkového svalového průřezu. Zajímavé je, že ve srovnání s řadou druhů zvířat je variabilita ve složení svalových vláken u člověka pomerně značná. Obvyklý průměr podílu rychlých vláken v zevní hlavě čtyřhlavého stehenního svalu (vastus lateralis) je kupříkladu kolem 55%, ovšem Simoneau a Bouchard (1989) zjistili u 418 mužů a žen rozpětí od 15 do 85%. Právě tyto rozdíly určují sportovní předpoklady každého jednotlivce. Sprinteři světové třídy mají například ve vastus lateralis až 70-90% rychlých svalových vláken, středotraťaři jen cca 30-50%. Naopak vytrvalostní běžci mohou mít přes 90% vláken typu I.
Vyšší procento rychlých vláken je často spjato s vyšší cirkulací tělesného testosteronu. Je známo, že atleti v rychlostních a silových sportech mají vyšší krevní koncentrace testosteronu než vytrvalci; hráči kolektivních her s charakterem přerušované aktivity (např. fotbalisté) se nacházejí zhruba uprostřed. Po 30. roce věku koncentrace tělesného testosteronu klesají a v souvislosti s tím se průřez rychlých vláken začíná pozvolna zmenšovat. Nakonec dochází u velké části z nich ke konverzi na vlákna pomalá. Důsledkem těchto změn je neodvratitelný pokles rychlosti a výbušnosti. Po 40. roce života dochází k atrofii obou typů svalových vláken, což má za následek pokles statické síly.
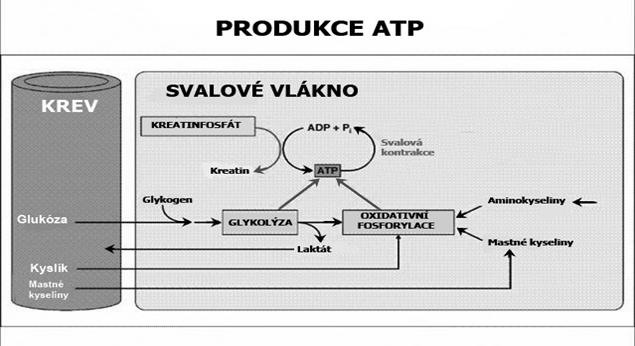
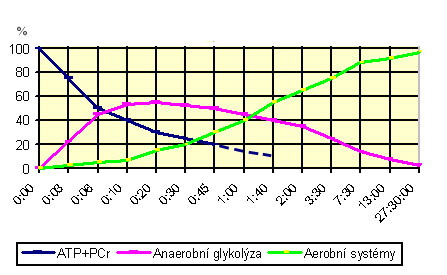
Energetické systémy lidského těla Svaly získávají ATP na pohyb příčných můstků prostřednictvím tří základních energetických reakcí: 1. Regenerací ATP z kreatinfosfátu („ATP-CP systém“) 2. Anaerobní glykolýzou („LA-systém“) 3. Aerobní oxidací glukózy a tuků („O2 systém“)
ATP+kreatinfosfát („ATP-CP systém“). Krátkodobé intenzivní výkony (sprint, vzpírání těžkých vah) je možno po omezenou dobu provádět anaerobně, tj. bez přístupu kyslíku. Během prvních sekund svalové práce je nejprve energie pro pohyb čerpána rozkladem malých zásob ATP uložených ve svalu. Když jsou tyto zásoby vyčerpány, je nový ATP regenerován reakcí ADP s kreatinfosfátem (fosfokreatinem), uloženým ve svalech. Z kreatinfosfátu se uvolní molekula organického fosforu a spojením s ADP vznikne nová molekula ATP. Tyto reakce jsou dominantním zdrojem energie po dobu prvních cca 5-6 sekund a na rozdíl od anaerobní glykolýzy při nich nevzniká laktát. Během delšího cvičení se kreatinfosfát nestačí regenerovat a jeho podíl na celkové energetické produkci prudce klesá (při 6 sec. práci cca 50%, ale při 30 sec. už sotva 30%; viz review Bangsbo 1998). Po skončení zátěže se jeho zásoby ve svalech opět rychle obnoví (75-80% během cca 1 minuty, 100% během cca 2-3 minut). Z uvedeného plyne, že čím vyšší budou zásoby kreatinfosfátu ve svalech, tím déle a s větší energií bude možno provádět krátkodobý, vysoce intenzivní anaerobní výkon. Protože kreatinfosfát vzniká sloučením organického fosforu s kreatinem, využívá se umělý příjem vysokých množství kreatinu pro zvýšení zásob kreatinfosfátu a tím i pro zlepšení výkonnosti v disciplínách jako sprint či vzpírání.
Anaerobní glykolýza („Lactic acid/LA-systém“) čili anaerobní rozklad glukózy se rozjíždí pouze s malým zpožděním po ATP-CP systému a už po cca 6 sekundách se podíl obou systémů vyrovnává. Glukóza je nejprve rozkládána na pyruvát a ten je poté bez přístupu kyslíku odbourán na kyselinu mléčnou, z níž následně vzniká laktát (kyselina mléčná > laktát + H+). Pokud je glukóza získávána ze svalového glykogenu, čistý zisk anaerobní glykolýzy představují 3 molekuly ATP na 1 molekulu glukózy. Pokud však výkon trvá déle a glukóza je do svalu přiváděna také krví z jater, čistý výtěžek se sníží na 2 molekuly ATP, protože 1 molekula ATP je použita na chemickou úpravu glukózy v játrech. Při produkci energie anaerobní glykolýzou se ve svalu hromadí laktát, a to i přes jeho rychlé vyplavování do krve a další metabolizaci v játrech a ledvinách (tam je použit při syntéze glukózy), nepracujících svalech, či v srdci (přeměněn na pyruvát). Když dosáhne hladina laktátu určité úrovně, disociované ionty vodíku (H+) způsobí takový pokles pH, že dojde k inhibici enzymů a tím i svalových funkcí. Zvyšující se okyselování způsobuje dráždění nervových zakončení a známý pocit „pálení“. Rychlost poklesu pH závisí na pufrovací kapacitě krve a svalů, tj. na schopnosti neutralizovat volné vodíkové ionty a zpomalovat okyselování. Pufrovací kapacita má velký význam pro sportovní výkon a lze ji zvýšit tréninkem. Mezi hlavní pufry patří fosfáty, některé bílkoviny (hemoglobin, histidin) a zejména karboxylové kyseliny a jejich soli (HCO3‾, NaHCO3‾).
Anaerobní glykolýza je sice dosti neefektivní způsob získávání energie a asi dvakrát pomalejší než regenerace ATP z kreatinfosfátu, nicméně stále výrazně rychlejší než oxidace glukózy. Po cca 30 sekundách intenzivní práce jsou zásoby kreatinfosfátu prakticky vyčerpány a náhlé „najetí“ na pomalejší anaerobní glykolýzu zapříčiňuje známou „čtvrtkařskou krizi“ z důvodu snížené rychlosti produkce ATP a hromadění laktátu.
Aerobní oxidace glukózy a tuků („O2 systém“). Při výkonech trvajících déle než cca 60-70 sekund dominuje jako zdroj svalové energie oxidace glukózy (tj. štěpení glukózy za přítomnosti kyslíku). V cytoplazmě svalové buňky je nejprve glukóza rozkládána na pyruvát, jenž je následně metabolizován v mitochondriích v tzv. Krebsově cyklu (cyklu kyseliny citrónové). Tato závěrečná reakce (oxidativní fosforylace) vede ke vzniku vody (H20), oxidu uhličitého (CO2) a velkého množství energie (38 ATP). Když se výkon stupňuje, množství mitochondrií, oxidativních enzymů a přijímaného kyslíku nestačí odbourávat pyruvát a dochází k jeho přeměně na laktát v procesu anaerobní glykolýzy. Pokud jsou po cca 90 minutách intenzivního výkonu zcela vyčerpány zásoby glykogenu a krevní glukóza nepostačuje, svaly začnou využívat energii převážně oxidací tuků (resp. volných mastných kyselin ze zásobního triacylglycerolu) v Krebsově cyklu na H2O a CO2. Tento způsob produkce energie (lipolýza) nevytváří laktát, ale je méně ekonomický nežli rozklad glukózy, neboť na stejné množství energie vyžaduje asi o 7% více kyslíku. To nevyhnutně vyvolává vyšší požadavky na dodávku kyslíku (zvýšení ventilace) a protékání krve (vyšší srdeční výkon). Při produkci energie z tuků již nelze udržet vysoké pracovní tempo. Vyčerpání glykogenových zásob a „najetí“ na pomalejší oxidaci tuků se projevuje notoricky známou krizí po 30 kilometrech maratonského závodu (tzv. hypoglykémie). Vzhledem k obrovskému množství energie uloženému v tucích by teoreticky bylo možné vykonávat svalovou práci na oxidaci tuků téměř do nekonečna; prakticky to však není reálné vlivem dehydratace, narušení osmotické rovnováhy v tělesných tekutinách, přehřátí apod. Jako zdroj energie slouží v případě extrémní dlouhodobé zátěže i proteiny, a to hlavně tzv. větvené aminokyseliny (BCAA).
Po skončení výkonu je pomocí zvýšené ventilace zoxidován nahromaděný laktát a jsou doplněny energetické rezervy (ATP, kreatinfosfát). Tato zvýšená spotřeba kyslíku se běžně označuje jako „kyslíkový dluh“. Při výkonech delších než cca 30 sekund se laktát nahromadí takovým způsobem, že se prudce zvyšuje čas na zotavení. Běžci na 400 metrů proto potřebují k zopakování kvalitního výkonu mnohem delší odpočinek než sprinteři na 100 m.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||